St Anne’s Academic Review 9 – 2019
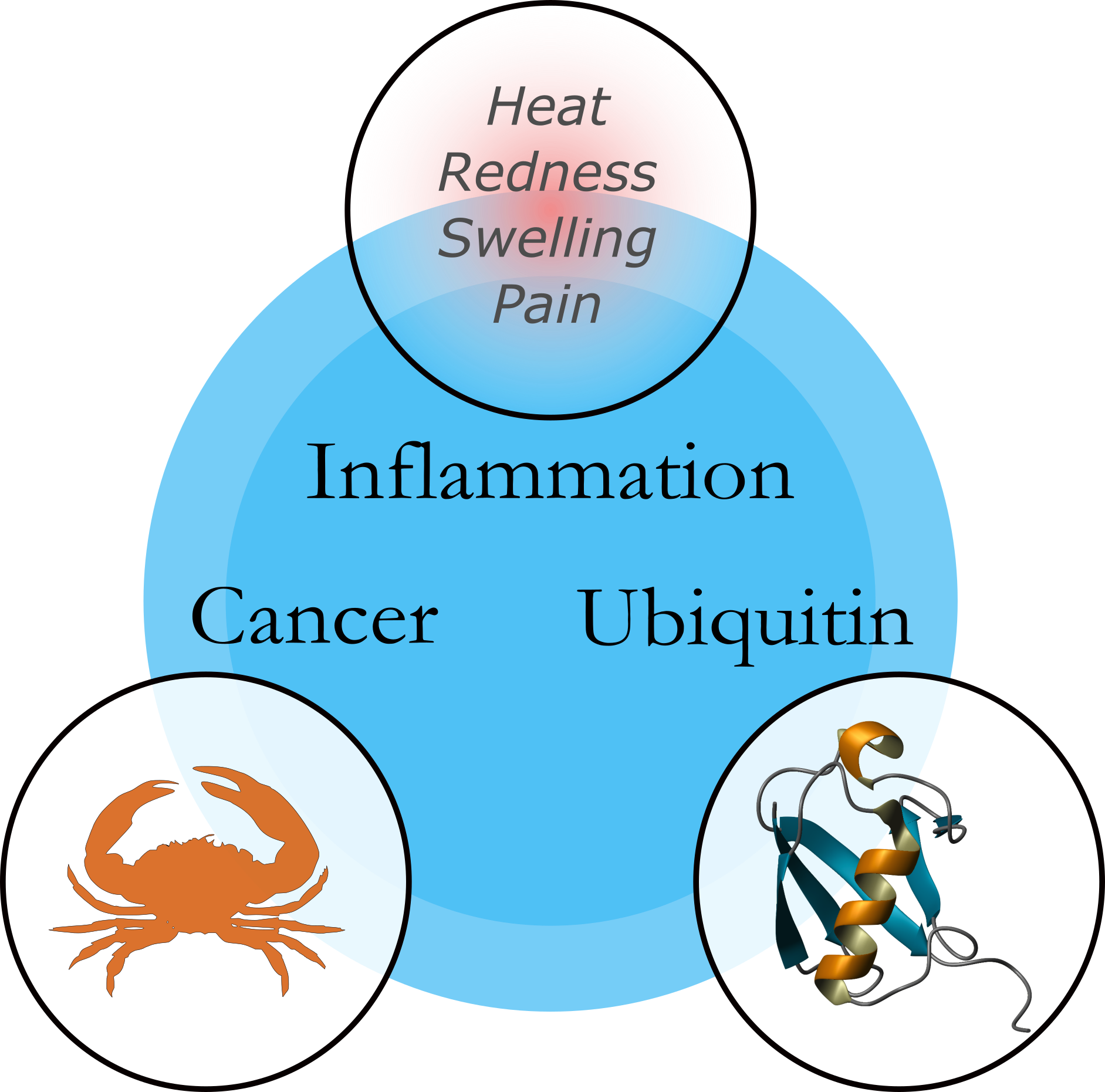
The molecular link between inflammation and cancer: ubiquitin
George Vere – MSc student, Department of Biochemistry
STAAR 9 – 2019, pp. 24-33
——————————–
Published: 13 October 2019
Review process: Open Peer Review
Draft First Uploaded: 25 July 2019. See draft and reviewers’ comments.
Abstract
Ubiquitin is a small protein central to the regulation of many cellular processes. Ubiquitination involves the covalent attachment of ubiquitin to other cellular proteins, and ubiquitin itself can be ubiquitinated to form polymeric chains. There are several sites on ubiquitin where other ubiquitin molecules can be attached, and as a result there are different ways that the ubiquitin chains can be assembled. The variety of chain types allows ubiquitin to play roles in many cellular processes, such as inflammation. When inflammatory molecules or pathogens are detected by a cell, ubiquitin chains are assembled within the cell on receptors and mediate a proinflammatory response by the cell. These non-degradative chains act as a scaffold to recruit and activate proteins required for activating NF-kB, the master regulator of inflammation. While inflammation is an essential part of the immune system and is normally a beneficial process contributing to the clearance of pathogens from tissues, deregulation of inflammation through chronic infection or mutation of proteins can lead to serious diseases, including immunodeficiencies and cancer. Inflammation can promote the survival and proliferation of cancerous cells. Mutations have been found which affect the assembly and disassembly of the ubiquitin chains in cancer patients, and recent studies have further suggested the importance of deregulated ubiquitin signalling in cancer. We have only just begun to understand the intricacies of ubiquitin in inflammatory signalling, and once it is better understood, it may present opportunities for therapeutic intervention in patients with inflammatory disorders or cancer.
Inflammation is an ancient concept in medicine, first described as redness and swelling with heat and pain in the first century A.D. by the Roman physician Cornelius Celsus [1]. Cancer is not a modern disease and has afflicted human civilisation for thousands of years. Bone cancer has been discovered in Egyptian mummies from around 2000 B.C.E., and the word cancer is derived from karkinos, the term used by Hippocrates to describe carcinomas and the Greek word for ’crab’, perhaps in reference to a crab lying in the sand, akin to a tumour spreading on the skin.
In the mid 19th century, the acceptance of cells as the building blocks of life and the proposal of the germ theory of disease hailed new era of understanding in biology [2]. Rudolph Virchow drew a direct link between inflammation and cancer in 1863, hypothesising that chronic inflammation was the cause of cancer, where irritants, tissue injury, and inflammation cause excessive proliferation (division) and growth of cells [1].
While we now understand that cancer involves many factors beyond cell proliferation, inflammation is key in providing an environment for tumours to develop. Solid tumours are complex structures, filled with immune cells, structural tissue, and dead tissue, possessing vastly altered metabolic profiles relative to normal tissue. Inflammation generates an interplay between the immune cells and cancer cells that both encourages cancer growth and suppresses the immune system [3].
In 2018, the Nobel Prize for Medicine or Physiology was awarded to James Allison and Tasuku Honjo for their pioneering work on cancer immunotherapy, which allows the harnessing of the immune system to kill cancer cells. Given that inflammation links cancer and immune cells, it has become more pressing than ever to discover how inflammation is regulated at the molecular level.
To understand the role of inflammation in cancer, we need to appreciate the normal role of inflammation in the body. Inflammation is one of the first defences of the body against invading pathogens. When a region of the body is infected by pathogen, it is recognised by the cells of the body, inducing an inflammatory response within the tissue which recruits other cells of the immune system to the area to fight the infection. This activates innate immune cells, which can immediately respond to the infection, and also primes an adaptive immune response, which require several days to come in to effect.
Cell signalling
Inflammation can, however, be damaging to cells and tissues, such that it must be limited, both in terms of its duration and the area of the body which is affected. As a result, tissues require a stimulus to mount an inflammatory response; a process which relies on cells being able to sense their environment. The stimuli come in the form of chemical messengers released by other cells to signal the need for an inflammatory response, or chemicals released by bacteria as they proliferate.
Cells must be able to respond to these inflammation-promoting molecules, and they do so via a process known as cell signalling. This serves to change the behaviour of the cells to generate an inflammatory response which involves the production of new proteins by the cell, which includes further inflammation-promoting molecules and proteins which alter the behaviour of the cell. However, the signalling process can also cause cells to die, a process generally referred to as cell death. The outcome of the signalling is complex and depends on the exact stimulus. At the heart of this signalling process lies the protein ubiquitin.
Inflammatory signalling
There are many extracellular factors which can initiate inflammatory signals in cells. The chemical signatures of pathogens, such as components of bacterial cell walls like lipopolysaccharide, are potent activators of inflammation. When our own cells sense these chemical messengers, they in turn release chemicals which affect other cells, causing an inflammatory cascade. The most important of these inflammatory molecules is tumour necrosis factor (TNF).
The power of TNF has been appreciated since it’s discovery in 1890, when the American surgical oncologist William Coley chose to research sarcomas following the death of one of his patients. He found the records of a surprising case, a patient who miraculously survived an apparently fatal cancer. Coley was convinced that the patient’s survival was due to contraction of a post-operative infection. This research lead to the discovery of ‘Coley’s toxin’, a bacterial extract injected into patients. The treatment, first trialled in 1896, allegedly gave some positive results, but the outcome is hard to judge as Coley did not systematically trial his treatment [4].
Throughout the 20th century, the mechanistic basis of Coley’s toxin was slowly unravelled. Lipopolysaccharide, a component of bacterial cell walls, was identified from this mixture in the 1940s, and was shown to induce tumour regression when isolated from bacterial extract [5]. However, as medicine advanced, more reliable and effective therapies, such as chemotherapy and radiotherapy, were developed and interest in Coley’s toxin decreased.
During the 1970s research on bacterial extracts resumed and new techniques in molecular biology enabled the identification of TNF as one of the agents responsible for the effects of Coley’s toxin [6]. However, TNF is not present in the bacterial extract itself – instead, the extract causes the host tissues to produce TNF and other inflammatory signalling molecules. TNF causes tumours to undergo necrosis, a form of cell death, and regress. It can cause cell death in certain cell types and cancer types, especially sarcomas. Injecting rodents with high doses of TNF caused tumour regression, as expected, but also had high toxicity.
In those with cancer, the injection of Coley’s toxin can cause increased activation of the immune system, leading to the selective targeting and destruction of the tumour. However, this is different from persistent and chronic inflammation, which can be beneficial for tumour survival. In this case, inflammation provides an environment which promotes cell proliferation, growth, angiogenesis (the growth of new blood vessels) and metastasis (the spread of cancer throughout the body). The variety of effects of TNF and inflammatory signalling shows that we need to understand the molecular basis of the signalling to determine why inflammation can promote cancer in some instances, but lead to tumour regression is others.
Ubiquitination
Inflammatory signalling in cells requires the protein ubiquitin. The name of ubiquitin betrays its identity – it is ubiquitously expressed by cells – but this tells us little about its function. Ubiquitin is a small protein, comprised of only 76 amino acids, in contrast to the hundreds of amino acids found in typical proteins. Its function is to modify other proteins, leading to changes in their behaviour, through covalent attachment to the protein; a process known as ubiquitination [7].
During this process, the end of ubiquitin is covalently attached to the target protein. The enzymes which carry out this process are known as E3 ligases. However, these enzymes do not work alone – they must be “charged” by the action of E1 and E2 ligases. This process is shown in Figure 1.
The addition of a single ubiquitin molecule to a protein is relatively common, but the real power of ubiquitination comes from the formation of polyubiquitin chains attached to a target protein. Once one ubiquitin molecule is attached to another protein, ubiquitin ligases catalyse ubiquitin polymerisation on the protein surface. Recent evidence has demonstrated that ubiquitin has eight different sites at which ubiquitin can be conjugated to itself by E3 ligases, enabling the formation of eight ubiquitin chains with different links between monomers [7].
Ubiquitin chains with alternative linkages have different shapes which are recognised by different binding proteins, with each protein partaking in different activities and roles within cells. Specific ubiquitin linkages can, therefore, perform different functions in the cell, the best studied of which is the targeting of a protein for destruction [7]. Ubiquitin chains tend to consist of one chain type, although branched chains containing different sub-types have been shown to exist [7], and are important in inflammatory signalling.
Of these eight ubiquitin chain types, two are important for mediating inflammatory signalling at the TNF receptor; K63-linked ubiquitin chains and linear ubiquitin. Ubiquitin chains can be trimmed down or completely removed from proteins by enzymes known as deubiquitinases (DUBs). This allows much better regulation of the length or presence of ubiquitin chains, analogous to the effect of the ‘brake’ in a car, playing a vital role in limiting the effect of ubiquitin ligases.
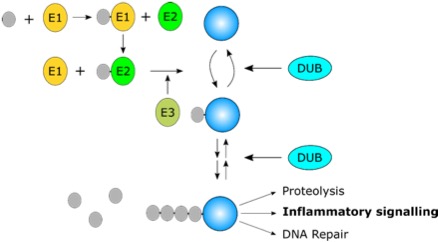
Figure 1: Ubiquitin (grey circles) chains are assembled on proteins (blue circle) by E1, E2 and E3 enzymes. Depending on the type of chain and the protein modified, ubiquitin chains lead to diverse outcomes, including inflammatory signalling. DUBs regulate the system by removing ubiquitin chains.
Ubiquitin in inflammatory signalling
The formation of ubiquitin chains is vital in inflammatory signalling. When TNF binds to its receptor on the cell surface (the TNF Receptor) it undergoes a change in shape leading to the association of many proteins with one another, forming a so-called receptor complex. Amongst these factors are E3 ligases, which polyubiquitinate other factors in the complex.
There is a hierarchical assembly of polyubiquitin chains at the TNF receptor complex. Initially, the E3 ligases assemble K63-linked ubiquitin chains. These serve as a scaffold to bring in another ubiquitin ligase known as the Linear Ubiquitin Chain Assembly Complex (LUBAC) to the receptor. LUBAC conjugates ubiquitin to various components of the TNF receptor complex, including to the existing K63-linked ubiquitin chains, and then on these ubiquitin molecules, assembles linear ubiquitin chains.
This leaves a complex ubiquitin landscapes, with both K63-linked ubiquitin chains and linear ubiquitin chains present at the receptor, and hybrid chains, that include both types of linkages. The linear ubiquitin chains are crucial for the following events in the signalling, and it still remains unclear why the branched chains are important [9].
Downstream signalling
The ubiquitin chains present at the receptor serves as a scaffold, to which other proteins bind and are activated. The further details in this signalling pathway are not important to our story, but the main effects of activation of these proteins is to enable the protein NF-kB to alter the proteins produced by the cell to promote further inflammation [10].
Inflammatory signalling can also cause the cell to adopt other fates. At the heart of this decision is the structure of the ubiquitin scaffold, which determines whether the cell will survive or die. Genetic alteration of cells has shown that in the absence of linear ubiquitin chains, cell death predominates. The location, timing and amplitude of the inflammatory signalling is altered by the structure of the ubiquitin scaffold. Further study will show exactly how the ubiquitin landscape shapes this decision [11].
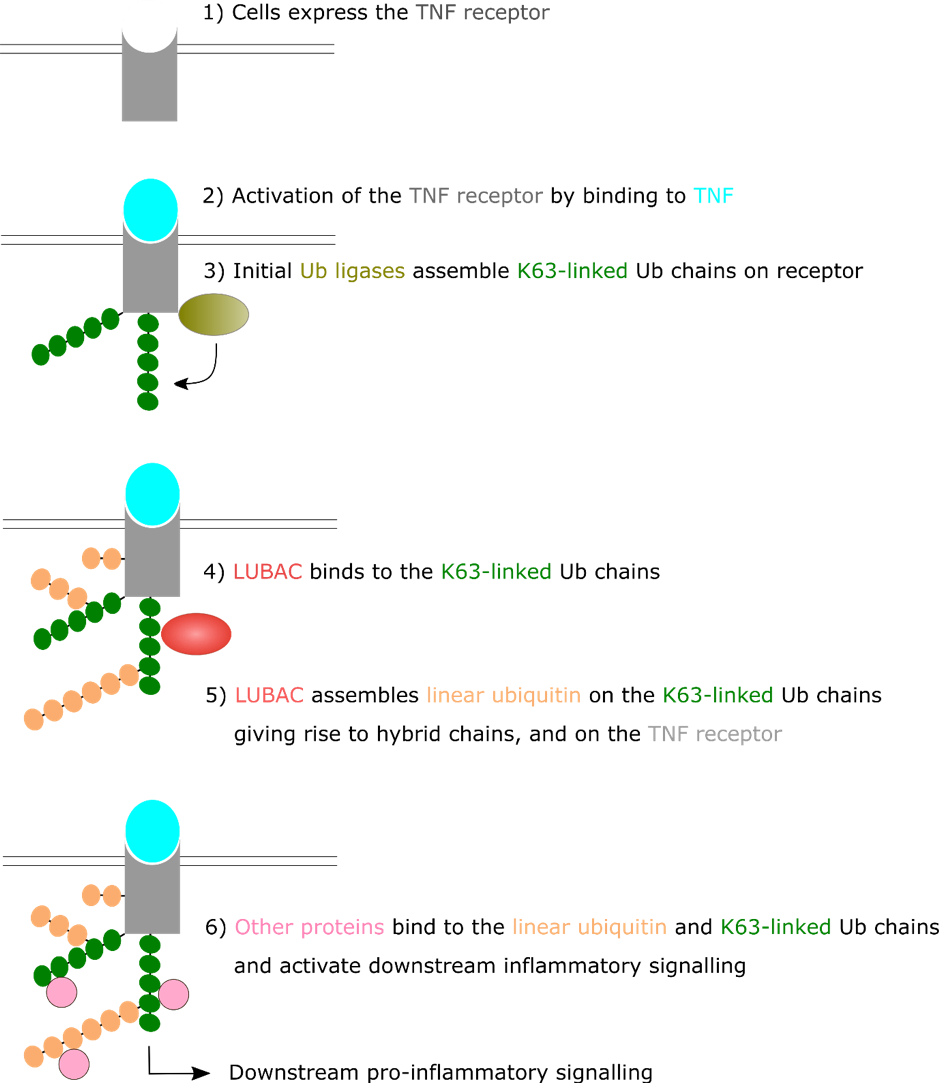
Figure 2: Tumour Necrosis Factor (TNF) activates the TNF Receptor causing a series of ubiquitination events that generates a complex ubiquitination landscape. The resulting ubiquitin chain landscapes tunes the downstream inflammatory signalling and can alter the fate of the cell.
Deregulation of ubiquitination
The length of polyubiquitin chains are tightly regulated by deubiquitinating enzymes (DUBs) capable of selective degradation of ubiquitin chains. In the case of inflammatory signalling, this allows the signal generated by the ubiquitin scaffold to be altered and fine-tuned. There are three DUBs which control ubiquitination on the TNFR complex, each of which has subtly different roles in editing the ubiquitin chain landscape and removing ubiquitin chains [12].
Ubiquitination and cancer
Deregulation of ubiquitination in inflammatory signalling results in several diseases, including chronic inflammatory conditions and cancer. Heredity diseases due to mutations in proteins which regulate TNFR complex ubiquitination are relatively rare but mice models in which components associated with linear ubiquitination and K63-linked chain formation have been genetically removed exhibit increased cancer incidence [9].
The hereditary disease cylindromatosis results in the formation of tumours in the head and neck and is caused by mutations in CYLD, one of the three DUBs that regulates TNFR ubiqutination. It not known exactly why mutations in CYLD lead to this disease, as CYLD functions in other inflammatory signalling cascades and other unrelated processes, including cell division (by regulation of microtubules), these mutations generally affect the catalytic activity of CYLD. CYLD activity is also associated with cell death [11]. In the case of TNF receptor signalling, CYLD tunes the inflammatory signal generated by the ubiquitin scaffold. The loss of CYLD in cylindromatosis could increase the ubiquitation of the TNF receptor, increasing inflammatory signalling and giving rise to an environment promotes tumour survival and proliferation.
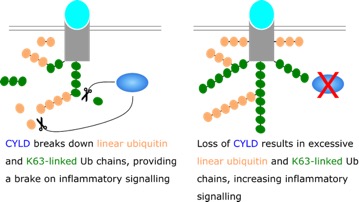
Figure 3: The loss of CYLD DUB activity causes excessive ubiquitination of the TNF receptor. The loss of CYLD is found in the cancer cylindromatosis, which could increase levels of inflammatory signalling by the TNF receptor.
Deregulation of linear ubiquitin chains has deleterious effects on cells. Loss of LUBAC causing excessive cell death in cell culture models [8]. In mice, genetic alteration of LUBAC components causes chronic skin inflammation. The loss of linear ubiquitin chains causes cell death in the skin of the mice, and the dead cells release inflammatory molecules into the extracellular environment. Cells can detect the molecules released by other cells as they die, and they trigger an inflammatory response. This positive feedback loop of cell death driving inflammation, driving cell death, causes chronic skin inflammation in the mice (Figure 4) [10]. While no direct link has been shown, chronic inflammation driven by loss of LUBAC could provide an environment for tumour survival and proliferation.
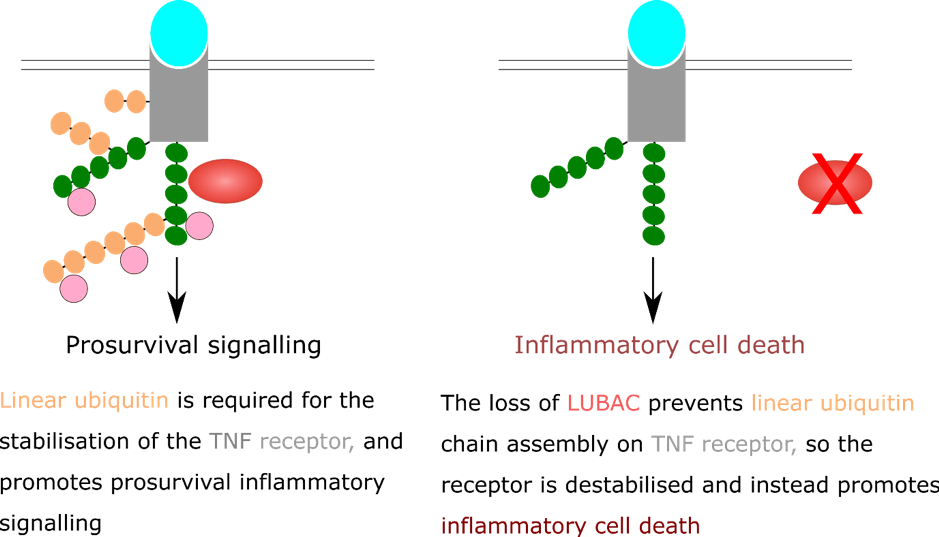
Figure 4: Without LUBAC, no linear ubiquitin chains are assembled at the TNF receptor. This activates a signalling pathway that results in cell death.
Summary and outlook
Inflammation affects tumour cells in opposing ways: it can either promote cell death as illustrated by Coley’s toxin but also provides an environment in which allows cancer cells to survive, as a result of TNF inducing pro-survival genes in cells. Linear and K63-linked ubiquitin chains are assembled at the TNFR complex following stimulation, and the ubiquitin chains coordinate events at the receptor required for either gene activation or cell death. However, much is left to be discovered about how the ubiquitin chains coordinate those events, and how the cell decides which fate to adopt.
Has our increased knowledge of ubiquitin’s role in inflammatory signalling given us tools for treating inflammatory diseases and cancer? While the degradative ubiquitin signal has been known about since the 1970s, the machinery that generates Linear ubiquitin was only discovered in 2006 so the machinery that modulates its assembly or breakdown is not yet targeted by any drugs [9, 10]. There are still many unanswered questions regarding the role of ubiquitin in inflammatory signalling, and this field will continue to grow. As we understand more about the interplay between cancer cells and inflammatory signals, we will be able to develop better therapies that can modulate the inflammatory signals in cells, allowing us to continue the War on Cancer into the 21st Century.
F. Rivas, “In this Issue: Inflammation,” Cell vol. 140, no. 6, pp. 755-757, 2010.
A. Sudhakar, “History of Cancer, Ancient and Modern Treatment Methods,” J Cancer Sci Ther, vol. 1, no. 2, pp. 1–4, 2010.
D. Hanahan and R. Weinberg, “Hallmarks of cancer: the next generation,” Cell, vol. 144, no. 5, pp. 646-74, 2011.
F. Balkwill, “Tumour necrosis factor and cancer,” Nature Reviews Cancer, vol. 9, pp. 361-371, 2009.
M. Shear and A. Parrault, “A Chemical treatment of tumors,” J Natl Cancer Inst, vol. 44, pp. 461-476, 1944.
Carswell EA Old LJ, Kassel RL, Green S, Fiore N, Williamson B., “An endotoxin-induced serum factor that causes necrosis of tumors.,” Proc Natl Acad Sci U S A., vol. 72, no. 9, pp. 3666-70, 1975.
D. Komander and K. N. Swatek, “Ubiquitin modifications,” Cell Res, vol. 4, no. 26, pp. 399-422, 2016.
Haas et al., “Recruitment of the Linear Ubiquitin Chain Assembly Complex Stabilizes the TNF-R1 Signaling Complex and Is Required for TNF-Mediated Gene Induction,” Molecular Cell, vol. 36, no. 5, pp. 831-844, 2009.
M. Hrdinka and M. Gyrd-Hansen, “The Met1-Linked Ubiquitin Machinery: Emerging Themes of (De)regulation,” Molecular Cell Review, vol. 68, no. 2, pp. 265-280, 2017.
H, Walzcak, “TNF and ubiquitin at the crossroads of gene activation, cell death, inflammation, and cancer,” Immunological Reviews, vol. 244, pp. 9-28, 2011.
D. Brenner, H. Blaser and T. W. Mak, “Regulation of tumour necrosis factor signalling: live or let die,” Nature Reviews Immunology, vol. 15, pp. 362-374, 2015.
M. Lork, K. Verhelst and R. Beyaert, “CYLD, A20 and OTULIN deubiquitinases in NF-κB signaling and cell death: so similar, yet so different,” Cell Death and Differentiation, vol. 24, pp. 1172–83, 2017.
Kirisako et al., “A ubiquitin ligase complex assembles linear polyubiquitin chains,” EMBO J, vol. 25, no. 20, pp. 4877-87, 2006.

The molecular link between inflammation and cancer: ubiquitin by George Vere is licensed under a Creative Commons Attribution 4.0 International License.
<< Back to Contents
<< Back to Publications
St Anne's Academic Review (STAAR) A Publication by St Anne's College Middle Common Room ISSN 2048-2566 (Online) ISSN 2515-6527 (Print)